
Project Report | March 20, 2020
Bonefish spawning in The Bahamas
Taking the Next Steps of Discovery
PI:
Aaron Adams, Ph.D. (Bonefish and Tarpon Trust; Florida Atlantic University, Harbor Branch Oceanographic Institute)
Co-Investigators:
Matthew Ajemian, Ph.D. (Florida Atlantic University, Harbor Branch Oceanographic Institute)
Sahar Mejri, Ph.D. (Florida Atlantic University, Harbor Branch Oceanographic Institute)
Paul Wills, Ph.D. (Florida Atlantic University, Harbor Branch Oceanographic Institute)
Jon Shenker, Ph.D. (Florida Institute of Technology)
Background
Worldwide, bonefishes (Albula spp.) are economically and ecologically important constituents of tropical, shallow-water systems. In the Caribbean Sea and western North Atlantic Ocean, Albula vulpes supports economically important recreational fisheries. For example, the estimated annual economic impact of the recreational fishery for bonefish, in The Bahamas, exceeds $169 million USD. Bonefish, tarpon (Megalops atlanticus), and permit (Trachinotus falcatus) comprise the recreational flats fishery that collectively generates an annual economic impact exceeding $465 million in the Florida Keys and $50 million in Belize. The economic importance of bonefish has led to the creation of strict regulations to protect the fishery in some countries: bonefish are catch and release only in Florida, Belize, Puerto Rico, and the U.S. Virgin Islands. In other locations, the recreational catch and release fishery coexists with consumptive fisheries that are subject to varying levels of management. In The Bahamas, capture with nets and commercial sale are illegal, but harvest with hook and line for personal consumption is allowed. The recreational flats fishery in Cuba occurs within marine protected areas designated as recreational catch and release zones, outside of these zones there are no regulations on what appears to be an intensive net fishery with high harvest. In addition to their fishery value, the abundance of bonefish in shallow coastal habitats, the dominance of benthic invertebrates in their diet, and their role as prey for sharks and barracudas suggest that bonefish play an important ecological role in structuring tropical shallow-water food webs. Recent and ongoing research has greatly increased our understanding of the bonefish life cycle. Adults exhibit high home range fidelity to shallow flats habitats of sand, seagrass, mangroves, and hardbottom during non-spawning time periods. Adults undergo migrations to pre-spawning aggregation (PSA) sites, which tend to be shallow protected bays near deep water, between October and April, with spawning mostly occurring near full moons. The PSAs move offshore at dusk to spawn at night in offshore waters before fish return to their shallow water flats habitats. After spawning, the eggs hatch in approximately 30 hours, and the larvae live as plankton for 41 – 71 days before metamorphosing into juveniles on sand or sandy mud bottoms in shallow, protected bays adjacent to deeper water channels that provide larval access and are near eventual adult habitats.
Conservation Needs
An International Union for the Conservation of Nature assessment classified bonefish as Near Threatened due to habitat loss and fragmentation (particularly mangroves and seagrasses), coastal development and urbanization, declines in water quality, and harvest by commercial, artisanal and recreational fisheries. In Cuba, for example, there is intense harvest of multiple bonefish pre-spawning aggregations (PSAs), with estimates of up to 20 tons harvested annually. In the Yucatan Peninsula of Mexico, fishers have traditionally targeted purported bonefish spawning migrations. In locations with recreational fisheries and no commercial harvest, the chief concern is habitat loss and degradation, though illegal harvest and lack of enforcement are also threats. For example, the top threats to bonefish in The Bahamas are habitat loss, habitat degradation, and illegal harvest of bonefish for commercial sale using nets. Habitat loss and degradation are caused by coastal development. Two of the PSA sites identified in The Bahamas, for example, have been proposed as sites for deepwater ports/marinas. Harvest is most common along spawning migration pathways. Threats to PSA sites are especially worrisome. Because individuals from a large geographic area are highly concentrated at PSA sites, these spawning aggregations and their associated populations are especially vulnerable to human impacts such as overfishing and habitat degradation and loss. Loss of productivity due to harvest or habitat disturbance at a localized aggregating site may have population-level consequences. For example, harvest of fish from spawning aggregations has caused regional population declines for Nassau grouper and other species, and coastal development has impacted Nassau grouper spawning sites in the Mexican Caribbean. Similarly, construction of causeways that disrupted bonefish spawning migrations on Kiribati have contributed to the cessation of spawning at numerous sites, and changes in bonefish population demographics. Recent oceanographic and genetic research revealed high levels of connectivity of bonefish populations among islands in The Bahamas as well as throughout the Caribbean. This means that when bonefish spawn, a significant portion of their larvae are transported to other locations. Thus, spawning at one site likely provides larvae to sustain the local population, but also supports different populations. For example, some of the larvae spawned on Abaco will settle on Grand Bahama and Andros. Thus, identification and protection of PSA sites has regional conservation implications. However, the oceanographic models that are used to estimate larval transport are based on surface currents, and treat the virtual larvae as passive particles in these currents. This is a significant shortcoming because the models do not include larval behavior and the depths at which they live, which might change estimates of larval transport.
Research Focus
The objectives of this research are threefold:
- Identify new bonefish pre-spawning aggregation (PSA) locations, and link them to home flats;
- Document and understand the offshore spawning movements of bonefish;
- Document and understand the reproductive physiology of female bonefish
- Document larval growth and behavior from hatching to metamorphosis.
Research Methods and Results
PSA Identification and Confirmation
Cruises in the 2019-2020 spawning season were to Abaco (November 2019), Crooked/Acklins (February 2020), and Bimini (March 2020). The PSA on Abaco is the most well studied and the PSA-home flats connection has been documented, therefore the focus of the research was on tracking and induction of spawning to study the reproductive physiology of females and the leptocephalus larvae.
Prior to the cruise to Crooked/Acklins we used Google Earth to select a list of potential PSA sites. We used flats boats, the Panga, and the Ribbed Inflatable to survey these sites extensively on multiple days during the week between the quarter and full moon during February 2020. No PSA was identified. During netting and tagging of bonefish on the flats, few bonefish exhibited characteristics indicative of pending spawning (large eggs, flowing milt). The extensive surveys allowed us to narrow the likely PSA site to three locations based on bathymetry, protection from weather, and connectivity to the flats via shallow migration pathways. PSA sites tend to be at the farthest up-current location from the flats that is accessible by near-continuous shallow water, is close in proximity to deep water, and is protected from prevailing winds. This will greatly reduce the amount of effort required to identify the PSA site in future work.
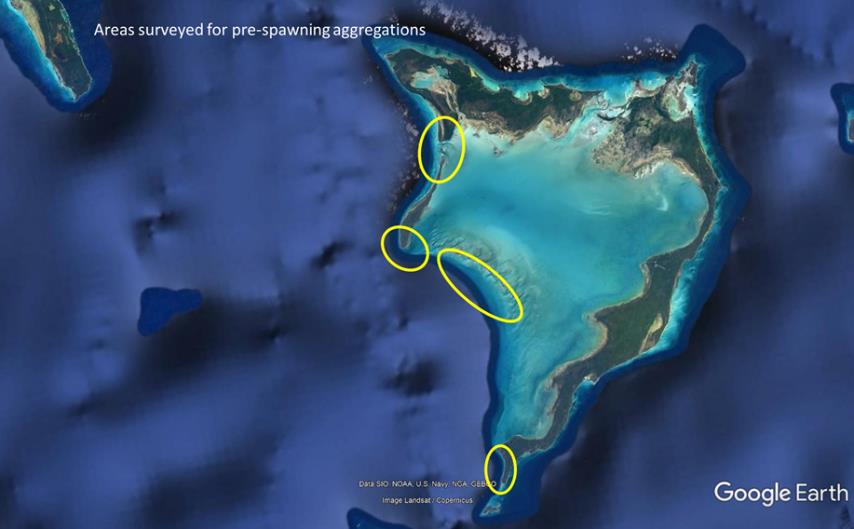
Areas of Crooked and Acklins surveyed for pre-spawning aggregation (PSA) sites during the spawning season of 2019-2020 (February 2020).
Approximately 70 miles of flats and channel habitats were surveyed throughout the Crooked Island and Acklins area during February 2020. A total of 299 bonefish was captured with a seine net, measured (fork length), tagged with dart tags, and released on the flats of Crooked and Acklins. Lengths ranged from 305 mm to 540 mm. One hundred twenty bonefish were evaluated for spawning readiness (i.e. sperm and/or eggs were extruded with gentle pressure on the abdomen). Overall, 9 males and 25 females were identified as ready to spawn. The absence of running sperm/eggs did not allow for sex determination of the remaining fish. Each tag has a unique identification number. Recaptures of these fish by local fishing guides and anglers will help identify the home ranges of these fish. It is also likely that fish will be recaptured during spawning migrations. When a PSA is found, tagged fish spotted in the PSA will be captured so that their home flat can be determined, thus identifying the range of influence of the PSA site.

The science team preparing to tag bonefish after capturing a school using a seine net during the February 2020 research cruise.

Bonefish being released after being dart tagged during the February 2020 research cruise.
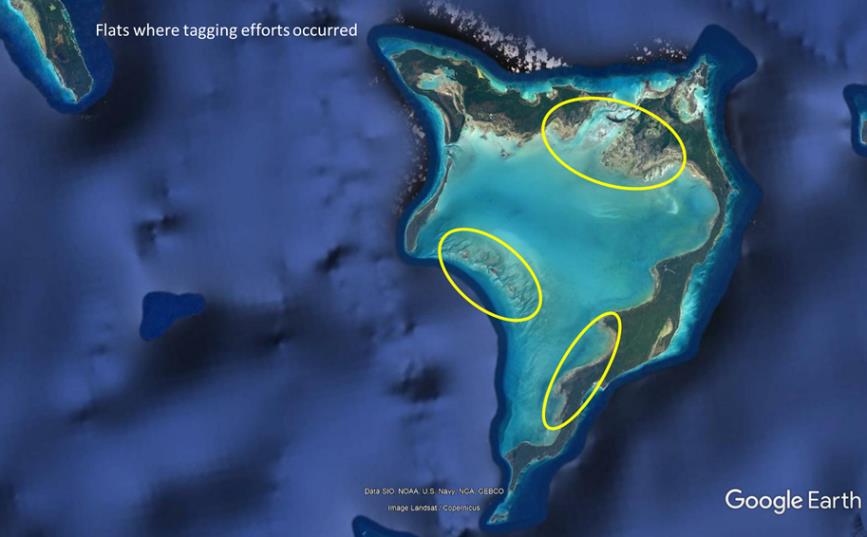
Flats on Crooked and Acklins islands, The Bahamas, where bonefish were captured with seine nets and tagged with dart tags during the February 2020 research cruise.
The PSA site first identified at Bimini in March 2019, was confirmed on the March 2020 cruise. It is notable that the locations where the PSAs were observed in March 2020 differed slightly from the sites identified in 2019. It is possible that the PSAs observed in 2019 were in transition to the PSA site identified this year, or were staging prior to moving to the 2020 PSA site. We have observed staging behavior at Abaco. Staging is when a school of fish migrating to the PSA site stops at a sheltered area for a day prior to continuing to the PSA site.
A total of 240 bonefish was tagged with dart tags on the flats. Size ranged from 342 mm to 560 mm. We identified 117 males, 33 females, and 55 were unidentified sex.
During the March 2020 cruise, we surgically implanted acoustic coded transmitters in four bonefish (two males and two females) captured from the PSA. These transmitters emit a ping at approximately two-minute intervals, and last for more than a year. The goal with these tags is to measure the rate of return to the PSA site to determine the number of times bonefish spawn in a season and estimate the peak month for spawning. We placed two acoustic receivers in the deeper basin adjacent to the flat where the PSA was observed to detect these tagged bonefish. The receivers will be retrieved in one year and the data downloaded.

Jewell Thompson (in the background), a colleague from Bahamas National Trust, working with the science team to tag and track bonefish. The Bahamas National Trust is a key partner in bonefish conservation since they designate and oversee national parks and protected areas.
Characterizing Offshore Spawning Movements
We surgically implanted continuous acoustic telemetry transmitters in bonefish at PSAs at Abaco and Bimini, and initiated tracks at each location. On Abaco, we implanted transmitters in four bonefish, two females and two males). Two of the fish appeared to be eaten by sharks, one tag failed. We were able to track the fourth tagged fish throughout the offshore spawning movement. The track at Abaco was the first complete track of bonefish offshore movement and spawning.
On 11 November 2019, one night before the full moon, southward offshore movements began at approximately 18:15 EST, 55 minutes after sunset. The fish followed the steep drop-off of the continental shelf as they migrated south, moving at depths of 0-5 m. Porpoising events, each lasting approximately 5 minutes, were noted four times during the offshore spawning migration.
Bonefish migrated a direct distance of 5.1 km from the PSA site to the location where they made the initial descent at 00:32 EST. The bonefish did not make a single descent to maximum depth, but made numerous descents and ascents. They reached a maximum depth of 136.7 m, and remained below 100 m for more than two hours. The spawning ascent was from 131.0 m to 67.3 m. The total time from leaving the PSA site to spawning was 14 hours 15 minutes. In addition to the telemetry track, we recorded the aggregation on boat-mounted sonar over the duration of the track, including the maximum depth recorded by the transmitter.

Spawning migration track from Abaco, November 11-12, 2019. The brown color at the top of the figure depicts the island shoreline. The blue background depicts the steep dropoff at the edge of the continental shelf. The red dots show the track of the bonefish school as it moves from shore to deep water, and then descends. We believe the final ascent before the track was terminated was the spawning event.
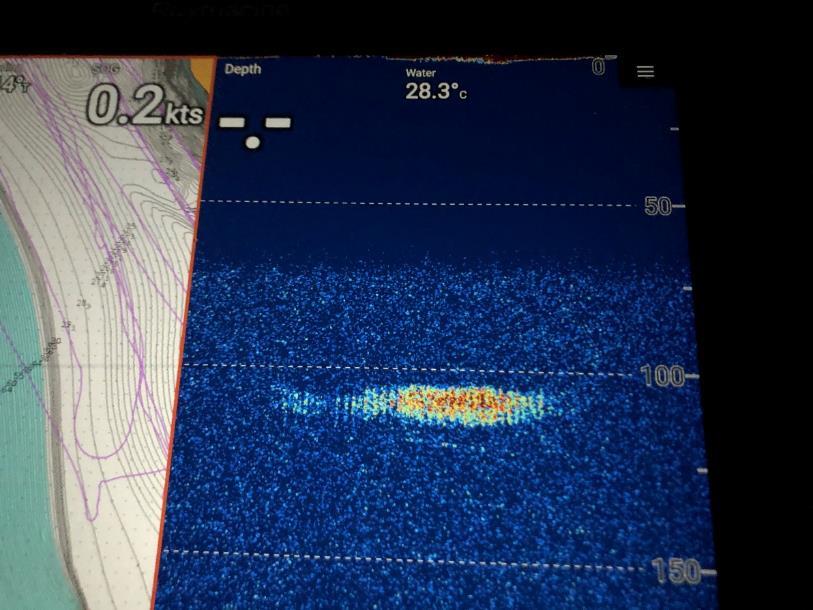
Sonar image showing the bonefish spawning aggregation at 110 m depth during the track of the spawning migration and spawning event.
Conductivity-Temperature-Depth (CTD) casts to a maximum depth of 100 m were taken each of the three days prior to the spawning run and during the spawning track using a Castaway CTD. These revealed an offshore pycnocline and thermocline, formations that were not present in waters less than 2.2 km from shore and inside the 200 m contour. The CTD cast at the location of the final spawning ascent found a pycnocline and thermocline at 75.9 m with measurements of 1024.0 kg·m-3 and 27.7 °C, respectively.

A CTD being deployed in offshore waters.
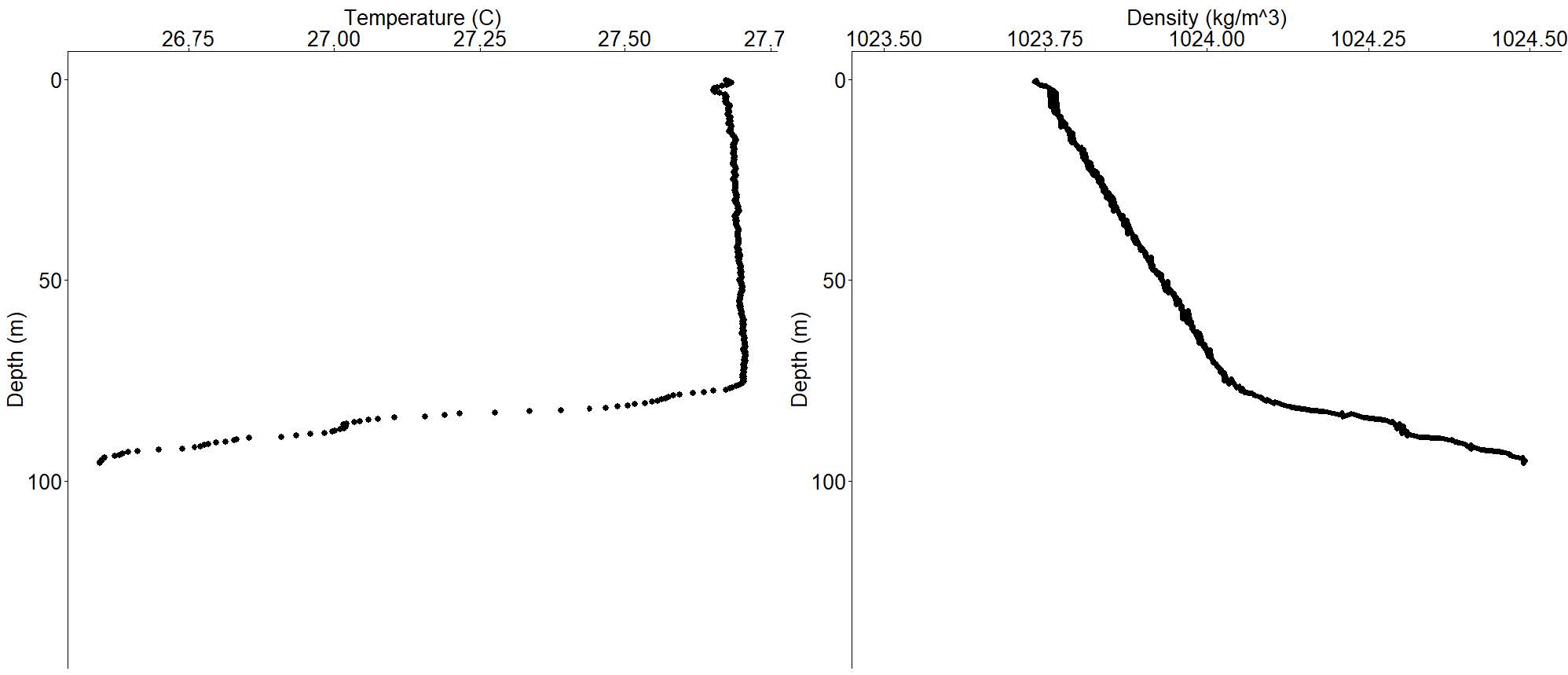
Graph showing the results of a representative CTD cast, showing the thermocline (abrupt change) in the left graph, and pycnocline (abrupt change) in the right graph at the spawning ascent location during the spawning track at the Abaco site.
We implanted continuous acoustic transmitters in six bonefish at the Bimini PSA site. On March 4, 2020, two fish were captured with hook and line from porposing PSAs in the basin adjacent to the flat where the PSA was observed, and fitted with transmitters. One fish appeared to be eaten by a predator (shark or barracuda). We tracked the other fish as it moved within the basin until 21:15 EST, at which time we aborted the track. This was likely a false run, which we have recorded at Abaco.
On the morning of March 5, in an attempt to re-locate the fish tagged on March 4we used the VR100 tracking equipment to survey the basin where tagging and tracking occurred. The tracking tags were not detected. We think both fish were lost to predation because on the morning of March 5, bonefish in the PSA on the flat showed signs of pre-spawning or spawning activity – many of the fish were bruised from the bumping and nudging that occurs during pre-spawning and spawning. The PSA on the morning of March 5 was approximately half the size of the previous day, suggesting either a high level of predation during the spawning run, or that a portion of the school returned to their home flats.
On March 5, we used a seine net to encircle the PSA on the flat, and implanted continuous acoustic transmitters in five female bonefish. The females were returned to the PSA, still encircled in the net. The net was then opened to free the PSA. The PSA remained on the flat until dusk. The PSA was monitored until approximately 20:00 EST, when the school appeared to split. We tracked one of the tags, and eventually determined it had been eaten by a shark. We then returned to the PSA site and found a second tag that also appeared to have been eaten by a predator. We visually checked the flat and discovered the aggregation was gone. Thus, as we were tracking fish that had been eaten by predators, the PSA had migrated off the flat, purportedly to spawn. We used the boat to make a number of transects searching for the remaining tags, but were unable to find the PSA. The track was terminated.

A pre-spawning aggregation (shown by the yellow arrow) at Bimini, The Bahamas, during March 2020.

Using the VR100 to track bonefish fitted with continuous acoustic transmitters at Bimini, The Bahamas during March 2020.
As on Abaco, we used a CTD to determine water characteristics offshore. Data from Abaco suggest that bonefish spawn at a depth where there is an abrupt change in temperature and density. The sampling at Bimini found a similar, though less strong, thermocline with a similar depth profile, but no change in density.
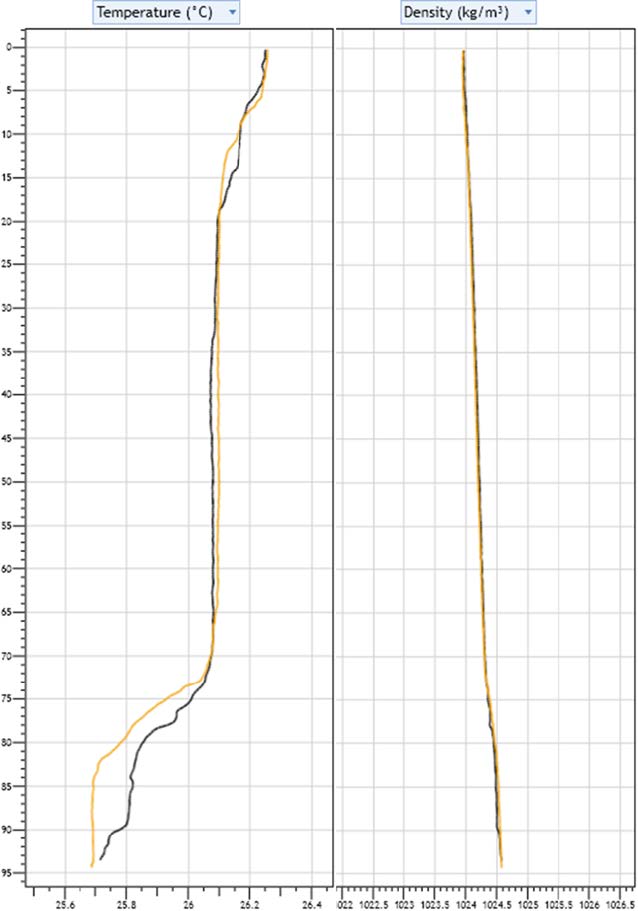
Graph showing the results of two representative CTD samples (orange and black) offshore of Bimini during March 2020. The change in temperature is similar in depth to what was observed in Abaco in November 2019. However, this was not sufficient to cause a pycnocline.
Conservation Implications
We have now identified or confirmed eight bonefish PSA sites in The Bahamas, two sites (Long Island and Bimini) within the past year, and have narrowed down the likely locations for two others (Crooked/Acklins and Berry Islands). Three of the PSA sites (two on Grand Bahama, one on Abaco) were already protected by designation as protected areas by Bahamas National Trust (BNT) in 2015.
Collaboration with Bahamas National Trust is essential to applying our research results to conservation. Bahamas National Trust is responsible for designating, operating, studying, and enforcing regulations for national parks and protected areas in The Bahamas. By collaborating directly with BNT on this research, we are able to share data directly with them. As important, the personal experience of BNT scientists collaborating on research excursions enables BNT to apply the data more effectively because they have understand the methods we use to study these important habitats and fishery. We will continue to work with BNT to create protections for the other PSA sites.
Because individuals from a large geographic area can be highly concentrated at aggregating sites, these spawning aggregations and their associated populations are especially vulnerable to human impacts such as overfishing, and
habitat degradation and loss. Important spawning aggregation sites have been effectively protected in some locations (e.g., red hind in the United States Virgin Islands). Indeed, as in this ongoing bonefish program, research can be conducted to identify spawning sites proactively – prior to habitat degradation – to inform conservation for important fisheries.
Habitat loss and degradation also threaten the functionality and long-term persistence of PSA sites. In some cases, PSAs have been identified in areas that are particularly vulnerable to development. For example, the proximity of deep water to a protected shoreline, an important characteristic for bonefish spawning, is appealing to those interested in developing deep-water ports or marinas. Indeed, a bonefish PSA site recently given national park protection in The Bahamas was previously targeted for development as a marina, and real estate developers have recently resubmitted a development proposal. Similarly, deep-water areas near ocean currents have been targeted for sewage outfalls. Therefore, BTT shares this information with non-governmental organizations, The Bahamas government agencies, and our fishery partners for incorporation into management strategies.
Like groupers that aggregate to spawn, bonefish likely locate spawning sites via social learning, whereby newly mature fish will follow older individuals to traditional spawning sites (traditional sites = used across generations). Under this scenario, habitat loss or degradation at a localized spawning site may result in population-level impacts. Individuals may continue to spawn at the traditional site, but alterations to the site may negatively affect reproductive success. If the PSA is able to shift to a non-traditional location following disturbance, important linkages between spawning site, larval transport pathways, juvenile habitats, and adult habitats may be severed. In previous work, we found that bonefish on Andros returned multiple times per year to the same PSA site. It will be interesting to learn the patterns of PSA site use from the four bonefish that were fitted with coded acoustic tags on Bimini.
The documentation of the offshore migration and descent to spawn during the Abaco cruise is not only new information for bonefish, but new to science. Never before has a species that lives in shallow water been documented descending to >130 m to spawn. Because the hydrostatic pressure is so much greater at depth than at the surface, many physiological processes are different, including blood chemistry and muscle physiology. The importance of these changes to the bonefish spawning process are unknown, and need to be studied.
While the Abaco spawning track is groundbreaking, additional tracks must be accomplished here as well as other locations to determine the level of variability in spawning behavior, better define the water characteristics that influence this behavior, and determine the impact of predators at different locations. For example, Bimini deserves additional tracking effort to determine the distance offshore and depth of spawning. Bimini is unique in that it is immediately adjacent to the Gulf Stream, where a two-knot current is common. In other locations where offshore spawning movements have been tracked (Abaco, Eleuthera), offshore currents are minor, and the shelf-break is closer to shore. If bonefish spawn far offshore of Bimini in the Gulf Stream, it is likely that their larvae are quickly transported northward into the open Atlantic, and would not contribute to the local bonefish population. This would mean that the bonefish population on Bimini is supported by larvae arriving from other locations. If, on the other hand, Bimini bonefish spawn at a location where a local current gyre occurs, the larvae might be retained in the area, or at least transported no farther than Grand Bahama, and thus contribute to the local bonefish population. This information is important to determining the level of protection that should be pursued for this location. In general, PSA sites that lose most of the larvae to long-distance transport into the Gulf Stream would be ranked lower in priority than sites that contribute larvae to bonefish populations.
Based on the track of the bonefish and the water profile data from the CTD on Abaco, it appears that bonefish were searching for the thermocline and pycnocline. Our hypothesis is that the bonefish spawn at the boundary layer to deposit the eggs, and thus larvae, in a region that provides appropriate food sources. The pycnocline boundary layer likely prevents the buoyant eggs from sinking, and can act to retain planktonic organisms, which includes prey to support larval growth. In addition, these deep boundary layers typically have low- turbulence and flow velocities, which may keep more larvae retained near the spawning location. These findings reveal that we must reassess our oceanographic larval transport models to accommodate the likelihood that bonefish larvae reside at depths rather than at the surface.
The new information on water characteristics at spawning depth is also helpful for better understanding conditions we need to emulate to successfully induce spawning, embryo development, hatching, and larval growth and survival. For example, it is likely that the enzymes essential to weakening the egg wall to allow hatching do not function well at higher temperatures. Similarly, it may be that a rapid reduction in water temperature as spawning is artificially induced may increase egg development and viability.
The 2019-2020 season built upon the large amount of new data collected during the 2018-2019 season, and resulted in numerous scientific firsts. Much of the data have already been applied to conservation efforts, and this will continue. Moreover, the findings on bonefish have broader conservation implications. The broad use of coastal habitats and offshore waters during the bonefish’s life cycle, the economic value of the catch and release recreational fishery, and the charismatic nature of the fishery make bonefish an appropriate umbrella species with broad conservation value in protected area designation. In this sense, umbrella species are “species whose requirements for persistence are believed to encapsulate those of an array of additional species.¨ Under the umbrella species concept, protected area network size and placement are set to protect habitats used by the single focal species, which offers protection for many other species that overlap in habitat use. Data on umbrella species therefore help to address the challenge of limited availability of data that are useful for marine protected area delineation and designation.